|
Arctic and Antarctica
Reference:
Karnysheva E.A., Filippova S.N., Brushkov A.V.
Examination of the ability of microorganism cells to migration in various dispersion frozen rocks
// Arctic and Antarctica.
2019. № 1.
P. 53-63.
DOI: 10.7256/2453-8922.2019.1.29226 URL: https://en.nbpublish.com/library_read_article.php?id=29226
Examination of the ability of microorganism cells to migration in various dispersion frozen rocks
Karnysheva Elina Andreevna
Post-graduate student, the department of Geocryology, M. V. Lomonosov Moscow State University
119234, Russia, g. Moscow, ul. Leninskie Gory, 1

|
Karnyshevaelina@mail.ru
|
|
 |
Filippova Svetlana Nikolaevna
PhD in Biology
Leading Scientific Associate, S. N. Vinogradsky Institute of Microbiology; Federal Research Center Fundamentals of Biotechnology of the Russian Academy of Sciences, Moscow
117312, Russia, g. Moscow, ul. Pr-T 60-Letiya oktyabrya, 7, korp.2
|
|
svfilipova@mail.ru
|
|
 |
|
Brushkov Anatoli Viktorovich
Doctor of Geology and Mineralogy
Professor, the department of Geocryology, M. V. Lomonosov Moscow State University
119234, Russia, g. Moscow, ul. Leninskie Gory, 1, of. 304
|
|
brouchkov@hotmail.com
|
|
 |
|
DOI: 10.7256/2453-8922.2019.1.29226
Received:
14-03-2019
Published:
21-04-2019
Abstract:
This article presents the results of model experiments on the ability of microorganism cells to migrate in pore space of the various dispersion frozen soils: Neogene sand of the Mammoth Mountain (Central Yakutia), quartz sand and kaolin clay at a fixed nedagive temperature (-6.5°C). Currently, there are known the basic parameters of microbial transport in the soils; however, the existing data on possibility of their migration in frozen soils and rocks is either contradictory or absent. The study was carried out on the example cells of Bacillus cereus strain F. The experiments were conducted in deep-freeze at a fixed negative temperature throughout one year. The question on the possibility of microorganism cells migration in permafrost is relevant from the perspective of the paramount global life preservation issues, stratigraphy of the various permafrost arrays, as well as their dynamics in the conditions of climate warming and emergence of risks of bringing microflora with the unknown properties to the modern ecosystems. Throughout the observation period, there was detected no evidence of migration of the model bacterial cells in frozen soils. The author established that the pattern of change in the number of colony-forming cells of the model organism in inoculated part of the samples of frozen soils is greatly affected by the level of their dispersion and aggregation.
Keywords:
frozen soil, permafrost, microorganisms, clay, sand, particle size, unfrozen water, metabolic activity, colony-forming unit, migration
This article written in Russian. You can find original text of the article here
.
Введение
Многочисленные исследования показали, что отрицательные температуры и стабильный физико-химический режим мерзлых осадочных пород благоприятствуют сохранению микроорганизмов [1, 9, 24, 25]. Это связано с незамерзшей влагой в мерзлых породах - основной эконишей, обеспечивающей сохранность клеточных структур в криобиосфере.
Изучение мерзлых толщ способствует пониманию состояния микроорганизмов, а микробиологические исследования, в свою очередь, полезны для характеристики мерзлых толщ. В литературе имеются данные о возможности проявления микроорганизмами вечной мерзлоты метаболической активности [16, 26, 28, 29, 30]. Перенос микроорганизмов в немерзлых почвах известен [6, 15, 17, 18, 19, 20, 21, 22, 23, 31], однако сведения о таком переносе в мерзлых почвах и породах противоречивы или отсутствуют. Изучение возможности движения клеток в мерзлых породах разной дисперсности может дать новую информацию о незамерзшей влаге в мерзлых породах (толщине, давлении, концентрации солей). Реальные размеры клеток (микроны), и расчетная толщина пленок воды (ангстремы), служащих эконишами для их сохранения, несопоставимы. Предполагается, что расчеты могут не отражать реальной картины распределения незамерзшей воды, представленной не только пленками, но и микрообъемными включениями [12]. Допуская, что микробные клетки не мигрируют и не делятся, мы можем считать их возраст равным длительности криоконсервации, то есть возрасту мерзлоты в синкриогенных толщах и времени последнего промерзания в эпикриогенных толщах. Самые древние мерзлые породы Арктики, из которых выделены микроорганизмы, датируются на северо-востоке Евразии поздним плиоценом (около 3 млн лет назад) [7].
Гранулометрический состав во многом определяет скорость миграции влаги и клеток микроорганизмов сквозь поровое пространство осадочных пород. Миграция микроорганизмов в грунтах зависит от комплекса физических и химических условий в системе. Выделяются 4 способа миграции в грунтах: движение в пленках воды вследствие самостоятельной подвижности микроорганизмов, рост гиф в пределах пленок воды вокруг минерального скелета, обеспечивающий движение от одной пленки к другой, рост микроорганизмов, способствующий миграции и дисперсия микроорганизмов, спровоцированная движением влаги в породе [27].
Есть данные по численности микрофлоры из сезонно-талого слоя почвы в зависимости от глубины, численность микроорганизмов остается достаточно высокой по всему почвенному профилю и накопление микроорганизмов происходит в нижних почвенных горизонтах, вплоть до границы с многолетней мерзлотой. Такой эффект авторы объясняют вымыванием веществ из верхних горизонтов и накоплением их над горизонтом мерзлоты [8].
Вопрос о возможности миграции клеток микроорганизмов в вечной мерзлоте является актуальным с точки зрения важнейших глобальных проблем сохранения жизни, стратиграфии различных слоев мерзлоты, а также их поведения в условиях потепления климата и возникновения рисков привнесения микрофлоры с неизвестными свойствами в современные экосистемы.
Целью настоящей работы было в условиях модельного эксперимента оценить возможность миграции клеток микроорганизмов в мерзлых породах разной дисперсности при отсутствии миграции влаги, то есть изучалось самостоятельное движение микроорганизмов в мерзлой породе при постоянной температуре.
Методы и материалы исследования
Модельный штамм (Bacillus cereus strain F), использованный в данном эксперименте был впервые выделен из мерзлых миоценовых отложений (песок неогенового возраста) в разрезе Мамонтовой горы (Центральная Якутия) [4] (рис. 1).

Рис. 1. Обнажение 50-метровой террасы реки Алдана. Фото Карнышевой Э.А.
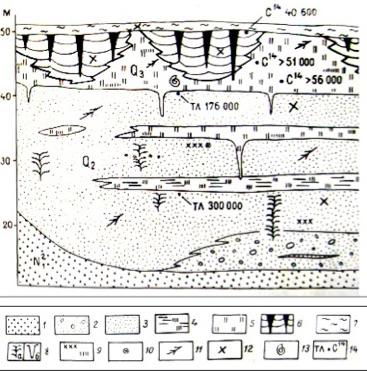
Рис. 2. Геологический разрез 50-метровой террасы Алдана в районе Мамонтовой горы: 1 – цоколь террасы: желтые косослоистые пески; 2 – галечники; 3 – пески серые разнозернистые; 4 – суглинки темно-серые заторфованные; 5 - суглинки сизовато-серые илистые; 6 - суглинки коричневато-серые с ледяными жилами; 7 - суглинки лёссовидные покровные; 8 – земляные жилы: отгибания (а), субаквального заполнения (б); 9 - линзы растительных остатков и детрита; 10 - шишки хвойных; 11 - обломки древесины; 12 - кости млекопитающих; 13 - раковины пресноводных моллюсков; 14 - термолюминисцентные (ТЛ) и радиоуглеродные (14С) датировки; 15 – номера слоев [11]
Мамонтова гора – геологически хорошо изученное и достоверно датированное обнажение реликтовых мерзлых толщ, простирающееся на 12 км вдоль левого берега р. Алдан в 325 км от его впадения в Лену. Представляет собой интенсивно размываемый речной эрозией останец водораздельной возвышенности Алдано-Амгинского междуречья, сложенный серией разновозрастных аллювиальных отложений видимой мощностью до 80 м. Формирование мерзлых толщ в данном регионе началось, вероятно, в позднем плиоцене около 3,5 млн. лет назад [3]. Позднее отложения, по-видимому, не оттаивали из-за холодного климата Якутии [2]. Ранее было проведено исследование изотопно-кислородного и дейтериевого состава позднеплейстоценовых и голоценовых сингенетических повторно-жильных льдов разрезов Мамонтова гора; Ю.К.Васильчуком в 1985 году было детально осмотрено все обнажение Мамонтовой Горы и прилегающих к нему низких террас и пойм Алдана [5].
Аэробная гетеротрофная бактерия, использованная в данном эксперименте, Bacillus sp. strain F, способна к росту в аэробных и анаэробных условиях; оптимальная температура роста - +37°C. По своей клеточной морфологии - это палочковидная, неподвижная, спорообразующая бактерия, размер клеток которой составляет 1-1.5 х 3 – 6 мкм (рис.3). Нуклеотидная последовательность гена 16S rRNA штамма F депонирована в DDBJ/EMBL/GeneBank под номером AB178889, идентификационный номер 20040510203204.24251 [4].
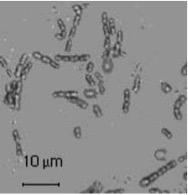
Рис. 3. Морфология клеток Bacillus sp. F. [4]
В качестве модельных грунтов использовали неогеновый песок, отобранный на Мамонтовой горе (Центральная Якутия) 1 м глубже слоя сезонного оттаивания вертикальных стенках речных обрывов, а также кварцевый песок (г. Люберцы, Московская область) и каолиновая глина (г. Новокаолиновый, Челябинская область) (Табл.1, 2).
Таблица 1. Геолого-генетическая характеристика грунтов
|
Номер №
|
Наименование породы
|
Возраст
|
Место отбора
|
Влажность, %
|
|
1
|
Песок неогеновый
|
N1
|
Мамонтова гора (Центральная Якутия)
|
17,9
|
|
2
|
Песок кварцевый
|
mJ3
|
г. Люберцы
|
19,7
|
|
3
|
Глина каолиновая
|
eP2
|
г. Новокаолиновый (Челябинская обл.)
|
40,7
|
Таблица 2. Гранулометрический и микроагрегатный состав грунтов
|
Наименование породы
|
Содержание частиц и микроагрегатный состав грунта, %
|
название грунта*
|
|
Диаметр частиц, мм
|
|
1 – 0,5
|
0,5 – 0,25
|
0,25 – 0,1
|
0,1 – 0,05
|
0,05 – 0,01
|
0,01 – 0,005
|
0,005 – 0,001
|
<0,001
|
|
Песок неогеновый
|
31,8
|
39,1
|
13,5
|
5,9
|
3,5
|
1,2
|
2,3
|
2,7
|
песок средней крупности
|
|
Песок кварцевый
|
6,5
|
6,5
|
79,6
|
2,2
|
2,4
|
0,4
|
0,3
|
2,1
|
песок мелкий
|
|
Глина каолиновая
|
0,7
|
0,5
|
0,4
|
2,9
|
19,5
|
11,2
|
40,2
|
24,6
|
глина пылеватая
|
*названия глин приведены, согласно классификации В.В. Охотина [14], песков – классификации Е.М. Сергеева [14].
Определение гранулометрического (зернового) и микроагрегатного состава проводили согласно принятым методам (ГОСТ 12536-2014): для глинистых грунтов– ареометрическим методом, для песчаных – ситовой метод с ареометрией осадка.
Для постановки модельных экспериментов использовали пробирки на ¾ объема заполненные образцами исследованных грунтов, которые затем стерилизовали при 121°C в течение 30 минут. Контроль стерильности производили путем высева образца на питательные среды). Последующие ¼ объема заполнялись соответствующим стерильным грунтом, инокулированным суточной культурой Bacillus sp. strain F. Конечная концентрация клеточной суспензии в соответствующих грунтах (т.е. сразу после внесения) составляла : для каолиновой глины КОЕ = 104 КОЕ/мл, для кварцевого песка КОЕ = 103 КОЕ/мл, для неогенового песка КОЕ = 103 КОЕ/мл.
Затем пробирки помещали в морозильную камеру, где поддерживали постоянную отрицательную температуру ( -6,5°С) (рис. 4).

Рис. 4. Пробирки с модельным грунтом (слева направо: кварцевый песок, неогеновый песок, каолиновая глина)
Эксперименты проводили по следующей схеме (рис. 5).
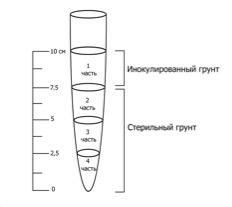
Рис. 5. Схема экспериментов
Наблюдения за наличием миграции бактериальных клеток с поверхностного слоя вглубь столбика грунта проводили через 1, 3, 6, 9 и 12 месяцев. Каждую инокулированную пробирку с грунтом распиливали при помощи стерильного напильника на 4 части (рис. 5).
Из каждой части производили высев на чашки Петри с питательной средой и инкубировали при комнатной температуре, после чего проводили подсчет колониеобразующих единиц и определяли значения КОЕ/мл. для каждой части образца в заданные сроки. Для культивирования микроорганизмов на чашках Петри использовали агаризованную питательную среду Agar Tryptose (Триптозный агар).
Влажность грунта на протяжении всего эксперимента оставалась неизменной, измерение влажности грунта проводили в сушильном шкафу при 105°С методом высушивания до постоянной массы, то есть влажность кварцевого песка оставалась ~ 19,7%, неогенового песка ~ 17,9%, каолиновой глины ~ 40,7%.
Было измерено содержание незамерзшей воды при температуре эксперимента (-6,5°С) контактным методом, для кварцевого песка оно составило ~ 0,11%, для неогенового песка ~ 1,32%, для каолиновой глины ~ 7,3%. Известно, что фазовый переход вода-лед в свободной воде происходит при 0°С, однако в пленках связанной структурированной воды он осуществляется при более низких отрицательных температурах и, чем более дисперсная порода, тем больше в ней содержание незамерзшей воды и тем тоньше пленка незамерзшей воды [14].
Результаты и обсуждение
В результате проведенных модельных экспериментов (табл. 3) не обнаружено наличие выраженной миграции клеток модельного микроорганизма с поверхностного слоя вглубь образцов исследованных типов грунтов. Предполагалось, что микроорганизмы будут мигрировать по прослоям незамерзшей воды, которые являются неотъемлемым элементом мерзлых пород. Однако, малые размеры (~ 0.01 - 0.1 микрон) пленок незамерзшей воды препятствуют свободному перемещению клеток.
Таблица 3.
|
длительность эксперимента
|
грунт |
часть образца |
каолиновая глина |
неогеновый песок |
кварцевый песок |
| 0 месяцев |
инокулированный |
1 |
104 |
103 |
103 |
| 1 месяц |
инокулированный |
1 |
102 |
101 |
101 |
| стерильный |
2 |
0 |
0 |
0 |
| 3 |
0 |
0 |
0 |
| 4 |
0 |
0 |
0 |
| 3 месяца |
инокулированный |
1 |
103 |
102 |
102 |
| стерильный |
2 |
0 |
0 |
0 |
| 3 |
0 |
0 |
0 |
| 4 |
0 |
0 |
0 |
| 6 месяца |
инокулированный |
1 |
101 |
102 |
103 |
| стерильный |
2 |
0 |
0 |
0 |
| 3 |
0 |
0 |
0 |
| 4 |
0 |
0 |
0 |
| 9 месяцев |
инокулированный |
1 |
0 |
103 |
102 |
| стерильный |
2 |
0 |
0 |
0 |
| 3 |
0 |
0 |
0 |
| 4 |
0 |
0 |
0 |
| 12 месяцев |
инокулированный |
1 |
101 |
102 |
102 |
| стерильный |
2 |
0 |
0 |
0 |
| 3 |
0 |
0 |
0 |
| 4 |
0 |
0 |
0 |
На риc. 6 представлено изменение показателей КОЕ модельного организма в инокулированной верхней части образцов исследованных типов грунтов в течение всего срока наблюдений.
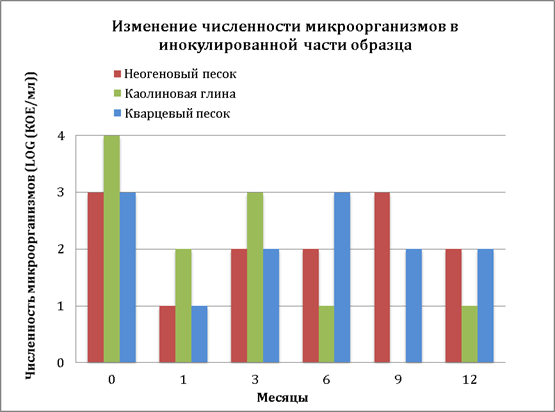
Рис. 6. Изменение численности микроорганизмов в инокулированной части образцов
В образцах песчаного грунта (кварцевый песок и неогеновый песок) изменение численности колониеобразующих клеток носило сходный характер. Резкое изменение условий окружающей среды при внесении клеточной суспензии в бедную питательными веществами среду и низких температур вызывали достаточно резкое падение численности КОЕ, отмеченное после 1 месяца наблюдений во всех типах грунтов. Необходимо отметить более низкий уровень падения численности КОЕ в образце каолиновых глин по сравнению с песчаными образцами. По мере адаптации к условиям среды наблюдалось небольшое увеличение количества КОЕ модельного организма. При этом, в песчаных грунтах увеличение показателя КОЕ происходило после 6 месяцев наблюдений, в то время как в образце каолиновых глин такое увеличение отмечено уже после 3 месяцев. Затем по мере увеличения срока наблюдений (до 12 месяцев) численность КОЕ в песчаных грунтах практически остается на одном уровне. В тоже время в образце каолиновых глин наблюдается постепенное снижение КОЕ модельного организма после 3 месяцев наблюдений и к концу 9 месяца колонии не обнаруживались. Отличительный характер изменения численности колониеобразующих клеток модельного организма в образце каолиновых глин можно объяснить процессами структурообразования в глинистых коллоидных системах [10, 13]. Указанные процессы с участием клеток микроорганизмов могут сопровождаться с течением времени изменениями состава электролитов в глинистых грунтах, их концентрацией, формированием коагуляционной структуры, характеризующейся присутствием тесных коагуляционных контактов, обуславливающих специфические свойства частиц осадка, которые могут осложнить процесс десорбции клеток с глинистых частиц. То есть глинистая коллоидная частица вместе с незамерзшей водой и адсорбированными ионами формирует сорбированный комплекс, в состав которых входят и адсорбированные на глинистых частицах клетки микроорганизмов, следствием которого может являться снижение способности к формированию КОЕ.
Выводы
1) Разработана методика проведения модельного эксперимента по изучению возможности миграции клеток микроорганизмов в мерзлых породах разной дисперсности с использованием в качестве модельного организма штамма Bacillus sp. F, выделенного ранее из образца отложений неогенового песка обнажения Мамонтовой горы (Якутия).
2) В результате экспериментов не обнаружено переноса клеток штамма Bacillus sp. F в исследованных образцах мерзлого песка и глины при -6,5°С.
3) Показано, что на характер изменения численности колониеобразующих клеток модельного организма большое влияние оказывает уровень дисперсности и агрегации грунта.
Благодарности
Авторы благодарны профессору Ю.К. Васильчуку за высказанные пожелания и замечания.
References
1. Abyzov S.S., Bobin N.E., Kudryashov B.B. Mikrobiologicheskie issledovaniya lednikovoi tolshchi tsentral'noi Antarktiki // Izvestiya AN SSSR, Seriya Biologicheskaya, 1979, № 6, s. 8–28.
2. Bakulina N.T., Spektor V.B. Rekonstruktsiya klimaticheskikh parametrov neogena Yakutii po palinologicheskim dannym // V knige: Klimat i merzlota, Pod red. G.N. Maksimova i A.N. Fedorova, Institut merzlotovedeniya, Yakutsk, 2000, c. 21-32.
3. Baranova Yu.P., Il'inskaya I.A., Nikitin V.P., Pneva G.N., Fradkina A.F., Shvareva N.Ya. Miotsen Mamontovoi gory // Trudy GIN SO AN SSSR, Moskva, Nauka, 1976, 376 c.
4. Brushkov A.V., Mel'nikov V.P., Griva G.I., Repin V.E., Brenner E.V., Tanaka M., Shchelchkova M.V. Biogeokhimiya merzlykh porod Tsentral'noi Yakutii // Kriosfera Zemli, 2011, tom 15, №4, s. 90 – 100.
5. Vasil'chuk Yu.K., Shmelev D.G., Budantseva N.A., Cherbunina M.Yu., Brushkov A.V., Vasil'chuk A.K., Chizhova Yu.N. Izotopno-kislorodnyi i deiterievyi sostav singeneticheskikh povtorno-zhil'nykh l'dov razrezov Mamontova Gora i Syrdakh i rekonstruktsiya pozdnepleistotsenovykh zimnikh temperatur Tsentral'noi Yakutii // Arktika i Antarktika. — 2017.-№ 2.-S. 112-135.
6. Devin B.A. Perenos mikroorganizmov v pochve i ego kolichestvennoe opisanie // Avtoreferat dissertatsii na soiskanie uchenoi stepeni kandidata biologicheskikh nauk, Moskva, 2004.
7. Demidov N.E., Gilichinskii D.A., Mironov V.A., Shmakova L.A. Kriobiosfera Zemli i poisk zhizni na Marse // Kriosfera Zemli, 2012, Tom XVI, № 4, s. 67-82.
8. Ivanova T.I., Kuz'mina N.P., Cherbunina M.Yu. Mikrobnoe soobshchestvo aktivnogo sloya pochvy iz obnazheniya Mamontovoi gory // Vestnik Severo-Vostochnogo nauchnogo tsentra Dal'nevostochnogo otdeleniya Rossiiskoi akademii nauk, 2017, № 4, s. 95–101.
9. Mel'nikov V.P., Rogov V.V., Kurchatova A.N., Brushkov A.V., Griva G.I. Raspredelenie mikroorganizmov v merzlykh gruntakh // Kriosfera Zemli, 2011, T. XV, № 4, s. 86–90.
10. Osipov V.I., Sokolov V.N., Rumyantseva N.A. Mikrostruktura glinistykh porod, Pod redaktsiei akademika E.M. Sergeeva // Moskva "NEDRA", 1989, 212 s.
11. Razrez noveishikh otlozhenii Mamontova Gora. Pod red. akad. K.K.Markova // Izdatel'stvovo Moskovskogo universitetata. 1973. 198 s.
12. Rogov V.V. Mikrostroenie merzlykh gruntov // Dissertatsiya na soiskanie uchenoi stepeni doktora geograficheskikh nauk, MGU, 1989, 337 s.
13. Sokolov V.N. Formirovanie mikrostruktury glinistykh porod // Sorosovskii obrazovatel'nyi zhurnal, 1998, № 7, s. 83–83.
14. Trofimov V.T., Korolev V.A., Voznesenskii E.A., Golodkovskaya G.A., Vasil'chuk Yu.K., Ziangirov R.S. Gruntovedenie // Pod red. V.T. Trofimova, 6-e izdanie, pererabotannoe i dopolnennoe, seriya "Klassicheskii universitetskii uchebnik", M., Izd-vo MGU i "Nauka", 2005, 1024 s.
15. Bai G., Brusseau M.L., Miller R.M. Influence of rhamnolipid biosurfactant on the transport of bacteria through a sandy soil // Applied and Environ, Microbiol., 1997, Vol. 63, p. 1866-1873.
16. Bakermans C., Tsapin A.I., Souza-Egipsy V., Gilichinsky D.A., Nealson K.H. Reproduction and metabolism at-10°C of bacteria isolated from Siberian permafrost // Environ Microbiol 5, 2003, p. 321–326.
17. Bales R.C., Li S., Yeh T.C.J., Lenczewski M.E., Gerba C.P. Bacteriophage and microsphere transport in saturated porous media: Forced-gradient experiment at Borden, Ontario // Water Resour. Res., 1997, Vol. 33, p. 639-648.
18. Baygents J.C., Glynn J.R., Albinger O. Variation of surface charge density in monoclonal bacterial populations: Implications for transport through porous media // Envrion. Sci. and Tech., 1998, Vol. 32, p. 1596-1603.
19. Camesano T.A., Logan B.E. Influence of fluid velocity and cell concentration on the transport of motile and nonmotile bacteria in porous media // Environ. Sci. and Tech., 1998, Vol. 32, p. 1699-1708.
20. Camper A.K., Hayes J.T., Sturman P.J., Jones W.L., Cunningham A.B. Effects of motility and adsorption rate coefficient on transport of bacteria through saturated porous media // Applied and Environ. Sci., 1993, Vol. 59, p. 3455-3462.
21. Chen K.C., Ford R.M., Cummings P.T. Mathematical models for motile bacterial transport in cylindrical tubes // J. Theor. Biol., 1998, Vol. 195, p. 481-504.
22. Corapcioglu M.Y., Haridas A. Transport and fate of microorganisms in porous media: A theoretical investigation // J. Hydrol., 1984. Vol. 72, p. 149-169.
23. Dahlquist, F.W., Lovely, P., Koshland, D.E. Quantitative analysis of bacterial migration in chemotaxis // Nature New Biol., 1972, Vol. 236, p. 120-123.
24. Gilichinsky D., Rivkina E., Shcherbakova V., Laurinavichuis K., Tiedje J. Supercooled water brines within permafrost—an unknown ecological niche for microorganisms: a model for astrobiology // Astrobiology 3, 2003, p. 331–341.
25. Gilichinsky D.A., Wilson G.S., Friedmann E.I. Microbial populations in Antarctic permafrost: biodiversity, state, age, and implication for astrobiology // Astrobiology, 2007, V. 7, p. 275-311.
26. Hultman J., Waldrop M.P., Mackelprang R., David M.M., McFarland J., Blazewicz S.J. Multi-omics of permafrost, active layer and thermokarst bog soil microbiomes // Nature 521, 2015, p. 208–212.
27. Parkinson D., Gray T.R.G., Williams S.T. Methods for studying the ecology of soil microorganisms // JBP Handbook No. 19, Blackwell Scientific Publications, Oxford and Edinburgh, 1971.
28. Rivkina E.M., Friedmann E.I., McKay C.P., Gilichinsky D.A. Metabolic activity of permafrost bacteria below the freezing point // Applied Environ Microbiol 66, 2000, p. 3230–3233.
29. Tuorto S.J., Darias P., McGuinness L.R., Panikov N., Zhang T., Häggblom M.M. Bacterial genome replication at subzero temperatures in permafrost // ISME J 8, 2014, p. 139–149.
30. Waldrop M.P., Wickland K.P., White R., Berhe A.A., Harden J.W., Romanovsky V.E. Molecular investigations into a globally important carbon pool: Survival strategies in ancient permafrost // The ISME Journal permafrost-protected carbon in Alaskan soils. Global Change Biology 16, 2010, p. 2543–2554.
31. Weiss, T.N., Mills A.L., Hornberger G.M. Effect of bacterial cell shape on transport of bacteria in porous media // Environmental Science and Technology, 1995, Vol. 29, p. 1737-1740.
|

 Eng
Eng













